Development
Reproduction and development are integral factors of life. Multicellular organisms arise through a process that begins with the fertilized egg and ends with a new individual. The fertilized egg undergoes cell divisions to increase the number of cells; simultaneously, the cells produced differentiate into the organs and organs systems of the fully formed organism. The scientific study of these developmental processes is called embryology. Aristotle (384–322 B.C.E. ), considered the first embryologist, described the growth of a chick embryo from a small dot of tissue to a fully formed bird.
Early Ideas in Embryology
Prior to the mid-1800s, scientists believed that development was the result of preformation. Preformation means that animals develop from an already existing miniature animal that merely required the right conditions to unfold and grow into a new organism. Scientific debates over whether the miniature animal was contained in the egg (ovum) or the sperm raged for decades. Scientists who believed that the miniature animal was in the egg were called ovists; those who believed that the miniature animal was in the sperm were called spermists.
In 1675, Marcello Malpighi (1628–1694), an ovist, reported seeing a miniature chick in the chicken egg. Anton Leeuwenhoek (1632–1723), a spermist, reported seeing a miniature human (homunculus) in each sperm (see Figure 1). In 1775, Lazzaro Spallanzani (1729–1799) demonstrated that both egg and seminal fluid were needed to produce a new individual. He conducted a series of experiments using amphibian eggs and seminal fluid. When the eggs were exposed to seminal fluid, they began to develop. However, if the eggs were exposed to filtered seminal fluid, fewer eggs developed. The more highly the seminal fluid was filtered, the fewer eggs developed. If the eggs were combined with the material left on the filter paper, they began to develop. Although Spallanzani correctly concluded that both egg and seminal fluid were necessary for development, he believed that the sperm seen in the seminal fluid were parasites . He postulated that the fertilizing agent was composed of the proteins and fats in the fluid.
William Harvey (1578–1657) viewed embryological development as a continuing process of remodeling and growth from unspecialized tissues to specialized structures. This theory, which is termed epigenesis, was largely ignored until 1759 when Kaspar Friedrich Wolff (1733–1794) offered empirical evidence of epigenesis from his detailed studies of chick development. In 1828, Karl von Baer (1792–1876) described four fundamental concepts of development. These four concepts, referred to as von Baer's law, are: (1) general features such as appendages appear earlier in the embryo than specialized features such as fingers; (2) development proceeds from general to specific characteristics; (3) as an embryo develops, it becomes increasingly different from other species; and (4) the early embryo of a higher animal is never like an adult of a lower animal, but only similar to the lower animal's embryo. Because they delineated the basic concepts of development, Wolff and von Baer are considered founders of modern embryology.
Fertilization
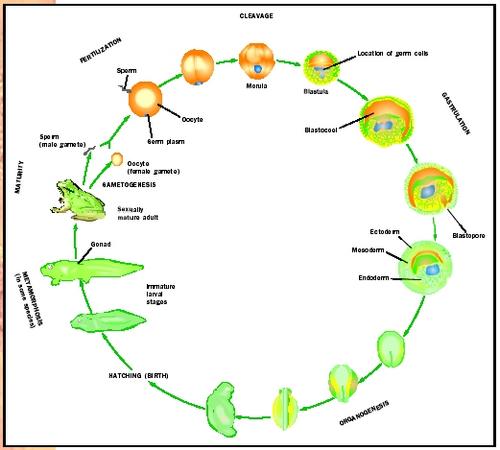
The process of development begins with the fusion of gametes : egg (ovum) and sperm. The motile sperm swims to the egg, pierces its cell membrane and enters the cell. Fertilization is the fusion of the nuclei of the egg and sperm, and the single cell that results from this fusion is called the fertilized egg or zygote (see Figure 2). During fertilization, the genetic material of the sperm and egg are combined. Each gamete is haploid , that is, it contains one-half of the normal number of chromosomes for the species. At fertilization, the gametes combine to produce a zygote with the full number of chromosomes for that particular species. Another way to say this is that fertilization restores the diploid number. For example, haploid human gametes have twenty-three chromosomes; when the egg and sperm fuse, the diploid state of forty-six chromosomes is restored to the zygote. Thus, each parent contributes one-half of the chromosomal complement of the new individual, resulting in a new organism with genetic characteristics of both parents. This single cell, the fertilized egg, gives rise to all the organs of the individual—muscles, brain, liver, eyes—through highly regulated and timed processes.
The stages from fertilization to the birth or hatching of an individual organism are identical to those of all individuals of the same species. Developmental stages include rapid cell division or cleavage; formation of the germ layers through the process of gastrulation; and differentiation and growth of the organs and organ systems. These stages are categorized as the periods of embryogenesis (cleavage and gastrulation) and organogenesis (formation of organs and organ systems).
Cleavage
As soon as the sperm enters the egg, the cell membrane of the egg undergoes changes that prevent the entrance of additional sperm. Meanwhile, the chromosomes from each parent come together and, within a few hours, the first cell division begins. The egg degrades the cytoplasm and organelles of the sperm; only the chromosomes of the sperm contribute to the fertilized egg.
The early cell divisions of the fertilized egg are called cleavage. The fertilized egg divides into two daughter cells called blastomeres. These two blastomeres divide into four blastomeres, the four blastomeres divide into eight, and so on. During cleavage, the total number of cells increases, but the size of each cell decreases. The reason for this strange situation is that cell division occurs so rapidly that there is not enough time for the individual cells to grow bigger. The constant doubling of cells during cleavage results in a multicellular embryo very quickly.
In a short period, the embryo has over one hundred cells arranged as a solid ball of blastomeres called a morula. The cells of the morula rearrange themselves into a single layer of cells surrounding a fluid-filled central cavity; the embryo at this stage is called a blastula (see Figure 2).
Gastrulation
The next step in development is the formation of the gastrula by invagination, the folding in of the cells of the blastula at a point called the blastopore. The resulting gastrula is a double-layer cup of cells. The outer layer of cells is termed the ectoderm and the inner layer of cells is termed the endoderm. The inner endodermal layer surrounds a new cavity, the primitive gut. A third layer of cells, the mesoderm, develops between the ectoderm and endoderm in most animals. Ectoderm, mesoderm and endoderm are the three germ layers from which all cells, tissues and organs develop (see Figure 2).
Cells of the ectoderm differentiate into the epidermis, hair, nails, claws, sweat glands, tooth enamel, brain, and spinal cord. Mesoderm differentiates into muscles, blood, blood vessels, heart, spleen, reproductive organs, and kidneys. Endoderm differentiates into the cells lining the digestive and respiratory systems, the liver, gallbladder, and pancreas.
Induction
One of the more fascinating aspects of development is the determination of body form, pattern, and differentiation. Put simply, how does a cell know what it is supposed to grow up to be? How do cells of the endoderm know they are supposed to form the digestive and respiratory systems? Induction is the process during which individual cells are "told" what they are supposed to become. A modern understanding of molecular events in development is discussed in the article Genetic Control of Development in volume 2 of this reference work. This essay outlines some pioneering work by Hans Spemann and Hilde Mangold.
Hans Spemann (1869–1941) received the Nobel Prize in 1935 for over twenty years of research on development in amphibians. In a series of elegant and delicate "baby hair loop" experiments, he demonstrated that when cells invaginate during gastrulation, they are induced to form specific cells and organs and that the primary inducer is a specific region of the blastopore. Spemann tied a strand of baby's hair around a fertilized newt egg so that the nucleus and some cytoplasm were on one side of the ligature while the other side contained only cytoplasm. After several cell divisions, Spemann loosened the ligature and allowed a nucleus to pass over into the other side. When cell divisions began on the side with the transported nucleus, the ligature was again tightened to separate the two masses of cells. The result was the production of two newt larvae, one a bit older than the other.

In later experiments, Spemann found that the location of the ligature was important. If the ligature were placed so that each half of the fertilized egg contained the certain cells (called the gray crescent because of their color) from the region destined to become the blastopore, two newts would develop. However, if the ligature were placed so that the gray crescent was only on one half of the cell, that part would form a newt, but the half without the gray crescent would remain a formless mass of cells he called the belly piece. Further experimentation demonstrated that during gastrulation cells became committed to their developmental fates.
Spemann and his graduate student Hilde Mangold (1898–1924) demonstrated that specific cells of the blastopore are the only determining region

Protostomes and Deuterostomes
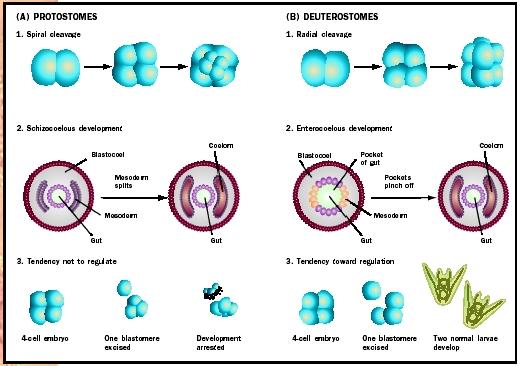
The fate of the blastopore is used to classify animals that have three germ layers into two large categories, protostomes and deuterostomes . Most adult animals have two external openings, the mouth and the anus, into the digestive tract. During gastrulation, the blastopore is the opening into the primitive gut. During further development, the blastopore becomes the mouth in animals classified as protostomes; the blastopore becomes the anus in deuterostomes. Organisms belonging to the phyla Mollusca (clams and snails), Arthropoda (insects and crustaceans) and Annelida (earthworms) are protostomes; members of the phyla Echinodermata (starfish) and Chordata (fish and humans) are deuterostomes. The type of cleavage and the development of the body cavity are other important differences between the protostomes and deuterostomes.
Protostomes exhibit spiral cleavage in which the blastomeres divide at acute angles to one another and are not aligned over one another. If one of these blastomeres is removed from the embryo, neither the removed blastomere nor the remaining cells develop into an individual. This type of determinate cleavage indicates that the fate of the daughter cells is determined early in development. A final characteristic of protostomes is that the body cavity, or coelom, develops as a split within the middle of the mesodermal layer. This type of coelom formation is termed schizocoelous development.
Deuterostomes, on the other hand, exhibit radial cleavage in which the blastomeres divide perpendicular or parallel to one another and are
Biogenetic Law
Ernst Haeckel (1834–1919), a physician, was so influenced by Charles Darwin's The Origin of Species that he gave up medicine and devoted himself to comparative anatomy. He disagreed with Darwin's theory of natural selection, and suggested that the environment acted directly on organisms, producing new species. In 1868, he proposed the biogenetic law, which sought to explain evolution as a series of stages in which the new characteristics of the next animal to evolve are simply added on to the lower animal. Briefly put, his biogenetic law stated that ontogeny recapitulates phylogeny (the
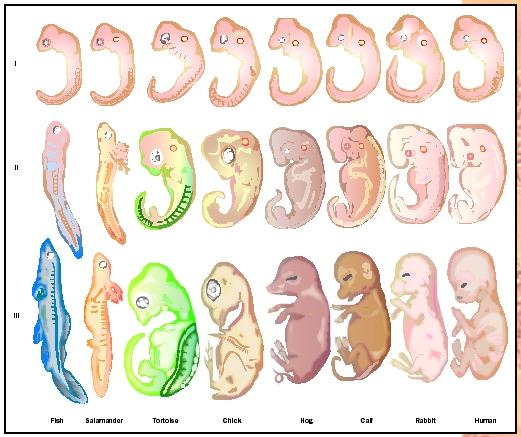
Modern scientists do not subscribe to the biogenetic law as postulated by Haeckel. However, there are elements of recapitulation that are important in comparative embryology. In 1828, Karl von Baer pointed out that vertebrates share common characteristics during development (see Fig. 3). Examination of vertebrate embryos reveals that during corresponding stages of early development, the embryos appear to be very similar. For example, all vertebrate embryos pass through stages in which they have gill pouches. The pouches eventually develop into the gill apparatus in fish; in later-evolving vertebrates that do not have gills, the gill pouches undergo further refinement and develop into structures associated with the head and neck. Similarly, all early vertebrate embryos have tails, which persist in some animals but regress during the later stages of development of humans. Thus, the individual development of an animal occurs through a series of stages that paint a broad picture of the evolutionary stages (phylogeny) of the species to which it belongs.
SEE ALSO Annelid ; Arthropod ; Clone ; Echinoderm ; Fetal Development, Human ; Genetic Control of Development ; Growth ; Meiosis ; Mollusk ; Reproductive Technology
Suzzette F. Chopin
Bibliography
Carlson, Bruce M. Human Embryology and Developmental Biology, 2nd ed. New York: Mosby, 1999.
——. Patten's Foundation of Embryology, 6th ed. New York: McGraw-Hill, Inc., 1996.
Gilbert, Scott F. Developmental Biology, 3rd ed. Sunderland, MA: Sinauer Associates, Inc., 1991.
Gould, Stephen J. Ontogeny and Phylogeny. Cambridge, MA: The Belknap Press, 1977.
Moore, Keith L., and T. V. N. Persaud. Before We Are Born: Essentials of Embryology and Birth Defects, 5th ed. Philadelphia, PA: W. B. Saunders, Co., 1998.