Cell Motility
Cells exhibit a wide range of movement. These movements include migration of cells along a surface or through a tissue, or movement of components within cells. Specific examples of cell motility include:
- • movement of cells from one location in an embryo to another during embryonic development
- • migration of cells into a wound during wound healing
- • contraction of a muscle cell that is the fundamental process responsible for muscle contraction
- • separation of a cell into two daughter cells during cell division
- • movement of membrane-bound vesicles into cells during phagocytosis or endocytosis
- • movement of membrane-bound vesicle from the cell interior to the cell surface during secretion
- • movement of chromosomes during mitosis
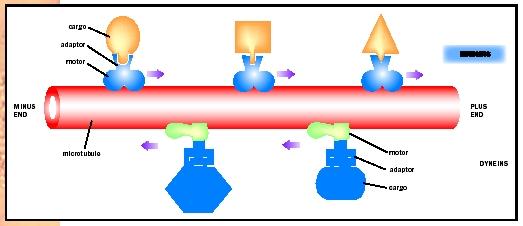
The first four bulleted points are examples of cell movement, while the last three bulleted points are examples of "intracellular motility." All of these movements have in common the fact that they are mediated by filamentous structures in the cell called the cytoskeleton and are powered by molecular motors that move along these filamentous structures. The simplest example is the movement of membrane-bound vesicles. These vesicles bind to a molecular motor just like a boxcar attaches to a railroad locomotive. The vesicle represents the cargo and the molecular motor represents the locomotive. The molecular motor then moves along the cytoskeletal filament as a locomotive moves along a railroad track. Most forms of intracellular movement occur using this mechanism.
A second mechanism is called contraction. This mechanism is responsible for contraction of muscle cells and the separation of daughter cells during cell division. Contraction works through the action of molecular motors pulling on the cytoskeletal filaments, drawing them toward each other. A third mechanism involves the rapid polyermization of the cytoskeleton. In this case the filamentous structures (usually the microfilament cytoskeleton) extend by the addition of subunits to the end. This growth of filaments then pushes out the membrane. This mechanism is responsible for protrusion of the front end of migrating cells.
Moving cells exhibit a special kind of directional movement called "chemotaxis." This mechanism accounts for the ability of cells to migrate in a specific direction. During chemotaxis, cells move in response to an external signal, most frequently a small molecule or short peptide, called a chemoattractant. Cells sense the concentration of the chemical and move in the direction of increasing concentration of the signal. This directional movement is responsible for much of the cell migration required for tissue formation and for wound healing. Wounded cells, for instance, release chemoattractants that attract immune system cells called macrophages and connective tissue cells called fibroblast .
For most eukaryotic cells , the process of cell movement occurs in several coordinated steps. First, cells extend a structure called a pseudopod ("false foot") using the polymerization mechanism. Next the pseudopod makes an attachment to the surface along which the cell is moving. This establishes the new front of the cell. A contraction-mediated process provides the force that moves the rest of the cell toward the front and leads to detachment of the trailing end of the cell. When cells are exhibiting chemotaxis they extend pseudopods in several directions, but only make the attachment to the surface in the direction of the highest concentration of the chemical signal.
In contrast, bacterial cells move by the action of an amazing rotary motor called a bacterial flagellum. This flagellum spins like a propeller propelling the cell forward. Bacteria undergo chemotaxis by a process called
SEE ALSO Connective Tissue ; Cytokinesis ; Cytoskeleton ; Endocytosis ; Exocytosis ; Immune Response ; Mitosis ; Muscle
Rex L. Chisholm
Bibliography
Alberts, Bruce, et al. The Molecular Biology of the Cell, 4th ed. New York: Garland Publishing, 2000.
Bray, Dennis. Cell Movements. New York: Garland Press, 1992.
Lodish, Harvey, et al. Molecular Cell Biology, 3rd ed. New York: Scientific American Books, 1995.
Comment about this article, ask questions, or add new information about this topic: