Chromosome Aberrations
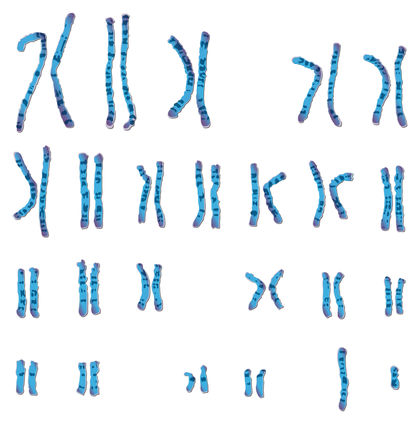
Chromosome aberrations are departures from the normal set of chromosomes either for an individual or from a species. They can refer to changes in the number of sets of chromosomes (ploidy), changes in the number of individual chromosomes (somy), or changes in appearance of individual chromosomes through mutation-induced rearrangements. They can be associated with genetic diseases or with species differences.
Trisomy
Chromosome number, size, and shape (X-shaped or V-shaped) are fixed for each species. In most animals, chromosomes are present in pairs, called homologous pairs, carrying similar genes. Each chromosome pair carries a distinctive set of genes. Genes code for proteins , and the amount of protein produced in a cell from a particular gene is proportional to the number of functional gene copies present.
Trisomy refers to having three copies of one chromosome. It arises through the chromosomal accident of nondisjunction during meiosis , which sends two copies of a particular chromosome into a sperm or egg, rather than one. An individual with a trisomy who survives to be born produces more of the gene products encoded on the trisomic chromosome. The resulting genic imbalance almost always severely impairs growth. Trisomy of the twenty-first chromosome, the smallest in humans, is the cause of Down syndrome, which is associated with mental retardation, congenital heart disease, accelerated aging, and characteristic facial features. Trisomy that occurs after fertilization , during fetal development, results in a "mosaic" individual with only some trisomic cells in the body. Such individuals may display some but not all of the features of the syndrome.
Trisomics for different chromosomes result in different abnormal characteristics. In humans, trisomies 13 and 18 are associated with different birth defects. While these individuals do not live long after birth, trisomies for any of the other non-sex chromosomes die before birth. Monosomies (having only one copy) for any chromosome also do not survive fetal existence, except for the sex chromosomes X and Y. Sex chromosome trisomies and monosomy for the X chromosome are associated with less severe effects on the phenotype .
Translocations
Translocations are the result of a chromosomal-level mutation, with two different (nonhomologous) chromosomes breaking and rejoining, placing the genes from one part of the one chromosome with part of the second chromosome, and vice versa. The number of genes is unchanged. Occasionally, the breakpoint mutation interrupts and inactivates the gene located at that chromosomal site. In other cases, the juxtaposition of new deoxyribonucleic acid (DNA) sequences from the other chromosome next to a gene at the breakpoint results in inappropriate expression. This action may activate an oncogene , for example. The "Philadelphia chromosome" is a translocation that fuses parts of chromosomes 9 and 22, which produces a new gene product that functions as an oncogene called Abl, which is implicated in chronic myelogenous leukemia.
Inherited translocations are passed through generations in a codominant fashion. Since one copy of each chromosome remains normal, both parent and progeny with such a translocation are heterozygous , or "balanced" carriers. Half their gametes will include one copy of each gene, either on the translocated chromosomes or their normal homologs. The other half, however, are unbalanced with some combination of translocated and normal homologs. The result is that the gamete has two copies of some genes, but no copies of other genes, from the translocated chromosomes. Such an "unbalanced" gamete, if it takes part in fertilization, often disrupts development so greatly that the individual does not survive to be born. If the number of unbalanced genes is low, however, children may be born, but often they have growth defects and mental retardation. Couples with recurrent spontaneous abortions may have one partner carrying a balanced translocation. Thus, gene copy number determines the specific phenotypes associated with a translocation, or with any chromosome aberration. Extreme examples of the importance of gene number are triploidy (3n 69 for humans) and tetraploidy (4n 92). These individuals nearly always die early in fetal life and are detected only in the remains of early spontaneous abortions. However, intolerance of polypoidy may be a mammalian phenomenon. It is common among plants, and frogs that are triploid are both viable and fertile.
Robertsonian translocations are a special class that result from the fusion of two V-shaped chromosomes at their centromere ends to form a single X-shaped chromosome. Individuals who are balanced for this translocation
have forty-five chromosomes, but are otherwise normal. However, during gamete formation, some gametes will become unbalanced, and their progeny are at risk for being aneuploid (without the correct set of genes). If the two fused chromosomes are homologs, then the risk is 100 percent that the zygote will be aneuploid since it will either have too few or too many genes. If nonhomologs are fused, the risk is usually 50 percent. About 5 percent of Down syndrome cases are caused by Robertsonian translocations.
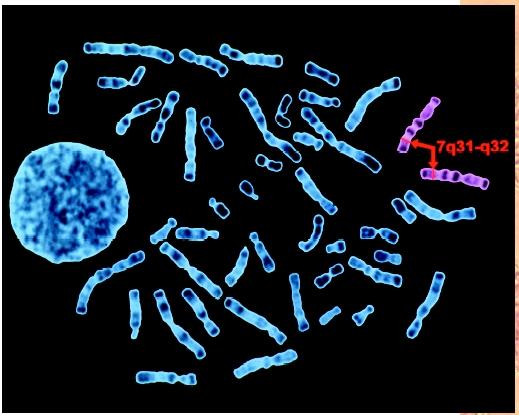
Chromosome fusions or exchanges at the centromere position can often be correlated with differences between closely related species. The great apes (chimpanzees and gorillas) have forty-eight chromosomes, for example. Their chromosome constitution differs from humans only in having one fewer small X-shaped chromosome pair and two additional small V-shaped pairs. At some point in our past, the V-shaped pairs fused at the centromeres to form the X-shaped pairs. Using DNA sequences that are conserved among vertebrate species as position markers, it is possible to create comparative maps of conserved blocks of genes. All mammalian X chromosomes are alike in the genes present, for instance. Thus, changes visible at the chromosome level are useful markers to follow the evolutionary relatedness of different species. Further comparison of conserved sequences suggests the vertebrate genome is a tetraploid (four-copy) version of the invertebrate genome.
Polyploidy
Changes in ploidy are of evolutionary importance in the flowering plants (angiosperms). Tetraploid plants often grow faster and larger than the diploid plants they derive from, and tend to be selected for agriculture. Alfalfa, coffee,
An example of plant polyploidy that affected human civilization and history is the origin of wheat. Modern bread wheat, cultivated for about eight thousand years, is a hexaploid with 2n 42, formed by sequential hybrids formed among three related grass species, each with 2n 14. To complete the circle, modern hybridizers have created a new species, Triticale, by crossing the ancestral Emmer wheat (2n 28) with rye (2n 14) and then doubling the chromosome number to 42 to take advantage of strong wheat growth with the high lysine content of rye. DNA analysis is scrutinizing the hybrid origins of many cultivated plants to identify their ancestors.
SEE ALSO Angiosperms ; Chromosome, Eukaryotic ; Gene ; Genetic Diseases ; Mutation ; Oncogenes and Cancer Cells ; Patterns of Inheritance
John Merriam
Bibliography
Gardner, R. J. McKinlay, and G. R. Sutherland. Chromosome Abnormalities and Genetic Counseling. Oxford: Oxford University Press, 1996.
John, Bernard. Meiosis. New York: Cambridge University Press, 1990.
Comment about this article, ask questions, or add new information about this topic: