RNA Processing
In the appropriate cell type and at the correct developmental stage, ribonucleic acid (RNA) polymerase transcribes an RNA copy of a gene, the primary transcript. However, the primary transcript may contain many more nucleotides than are needed to create the intended protein. In addition, the primary transcript is vulnerable to breakdown by RNA-degrading enzymes.
Before the primary transcript can be used to guide protein synthesis, it must be processed into a mature transcript, called messenger RNA (mRNA). This is especially true in eukaryotic cells . Processing events include protection of both ends of the transcript and removal of intervening nonprotein-coding regions.
On an RNA molecule, the end formed earliest is known as the 5 (5-prime) end, whereas the trailing end is the 3 end. The ends of the primary transcript are particularly susceptible to a class of degradative enzymes called exonucleases. During processing, the 5 end of the primary transcript is protected against the effects of these enzymes by the addition of a CAP. The CAP uses an unusual linkage between nucleotides. Exonucleases do not recognize this unusual structure and therefore cannot remove the CAP. Since exonucleases work only from an end, if the CAP nucleotide cannot be removed, the entire 5 end of the mRNA is protected. The 5 CAP also aids in transport out of the nucleus and helps bind the mRNA to the ribosome .
To protect the 3 end against degradative exonucleases, a poly-A tail is added by poly-A polymerase. Poly-A is a chain of adenine nucleotides, one hundred to two hundred units long. The poly-A tail has typical bonds that are susceptible to degradation by exonucleases, but it does not have any protein coding function so it does not particularly matter if some of the A
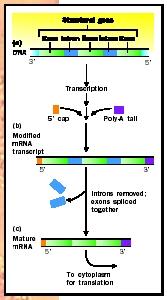
The most striking event in RNA processing occurs because the protein coding region in eukaryotic genes is not continuous. A typical eukaryotic gene is composed of a number of protein coding regions, called exons, that are separated by noncoding regions called introns. In fact, the number of nucleotides in the introns can be much larger than the number of nucleotides in the combined exons. The DNA gene contains the code for both the exons and the introns, as does the primary RNA transcript, but the noncoding intron sequences must be removed to form the mRNA before protein synthesis.
The process by which introns are removed and exons are joined to one another is called RNA splicing, and it is catalyzed by complexes of proteins and RNA called SNuRPs (small nuclear ribonucleoprotein particles). These complexes locate special RNA sequences that flank the exon/intron junctions, bind to them, and catalyze the splicing reactions. Some primary transcripts can be spliced in a few different ways. Such "alternate splicing" yields a range of related proteins.
After addition of the CAP to the 5 end, the poly-A tail to the 3 end, and splicing of the introns, the processing is complete and the mRNA is transported through nuclear pores to the cytoplasm of the eukaryotic cell where translation (protein synthesis) will occur.
SEE ALSO Gene ; Nuclear Transport ; Protein Synthesis ; RNA ; Transfer RNA ; Transcription
James E. Blankenship
Bibliography
Alberts, Bruce, et al. Molecular Biology of the Cell, 4th ed. New York: Garland Publishing, 2000.
Stryer, Lubert. Biochemistry, 4th ed. New York: W. H. Freeman and Company, 1995.
β-thalassemia, a hemoglobin disease, can be caused by an intron mutation that prevents recognition of a splice site.
Comment about this article, ask questions, or add new information about this topic: